Over at Sandwalk, Larry Moran posted some interesting bits rrom his molecular evolution class exam, including a passage from Mike Lynch arguing for his claim that “nothing in evolution makes sense except in the light of population genetics”. In this passage, which I’ll quote below, Lynch says that evolution is governed by 4 fundamental forces.
The idea that evolution is governed by population-genetic “forces” is common but fundamentally mistaken. I wish we could just put this to rest.
The entire passage from Lynch’s Origins of Genome Architecture is this:
Nothing in Evolution Makes Sense Except in the Light of Population Genetics
Evolution is a population genetic process governed by four fundamental forces, which jointly dictate the relative abilities of genotype variants to expand through a species. Darwin articulated a clear but informal description of one of those forces, selection (including natural and sexual selection), whose central role in the evolution of complex phenotypic traits is universally accepted, and for which an elaborate formal theory in terms of changing genotype frequencies now exists. The remaining three evolutionary forces, however, are non-adaptive in the sense that they are not the function of the fitness properties of individuals: mutation (broadly including insertions, deletions, and duplications) is the fundamental source of variation on which natural selection acts; recombination (including crossing-over and gene conversion) assorts variation within and among chromosomes; and random genetic drift insures that gene frequencies deviate a bit from generation to generation independently of other forces. Given the century of theoretical and empirical work devoted to the study of evolution, the only logical conclusion is that these four broad classes of mechanisms are, in fact, the only fundamental forces of evolution. Their relative intensity, directionality, and variation over time define the way in which evolution proceeds in a particular context.
I would like to disagree with every sentence. Let’s start with the first one.
Evolution is a population genetic process governed by four fundamental forces, which jointly dictate the relative abilities of genotype variants to expand through a species.
The “forces” analogy popularized by Dobzhansky comes from physics. It has been discussed extensively by philosophers since Elliot Sober promoted the idea that evolutionary theory is “a theory of forces” (for explanation, see the blog by Kele Cable). The idea is that we have a system defined by some dimensions, which in classical Newtonian physics are the dimensions of physical space, and there are independent “forces” that push objects around within this space. A force is a force because it has a generic capacity to displace a particle from A to B, or more properly, because it can alter the momentum of a particle independently of other forces. Forces have nice additive properties, so that the system doesn’t become complex and unpredictable if we have >1 force. We can just use vector addition to combine forces based on their magnitudes and directions (image from Wikipedia on vector addition). Because the forces all act independently, we can add a 3rd force to the resultant vector of 2 other forces, and we’ll still get the correct result. 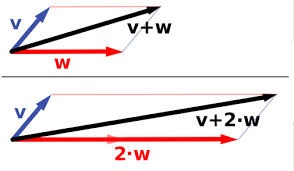
Population-genetics theory is like that in some ways. We can imagine a zero-force law for the system “at rest”, with perfect inheritance— that’s Hardy-Weinberg. We all know the 1-force model of allelic selection, with a nice sigmoidal curving showing an allele frequency going from low to high. We can make a 2-force model called the mutation-selection balance model, where there is just mutation pushing the frequency of deleterious allele A1 up, and selection pushing the frequency back down. The system reaches a fixed point, the equilibrium frequency, which is simply u/s (haploid case).
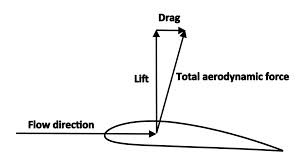
Yet the analogy breaks down very quickly, in important ways. Forces have (1) magnitude and (2) direction. Drift and recombination have magnitude but not direction, thus they are not forces. Furthermore, selection and drift are entangled, not independent, because they are just 2 artificially separated components of the same process, like separating aerodynamic force into lift and drag (image).
But those are tiny problems compared with the way the “forces” view fails to cover the introduction of a novel variant. Everyone knows that the space in which the forces operate is defined by allele frequencies. What few have noticed is that the forces analogy only makes sense if these are non-zero allele frequencies, i.e., the forces operate in the topological interior of an allele-frequency space.  This is the only place where the 4 forces have any kind of parity or independence: any force can shift f to f + d, where f is a frequency such that 0 < f < 1, and d is an infinitesimal. This is what makes a force a force. By contrast, if we are not in the interior— if we are sitting on an axis waiting for a new mutation—, then selection, drift and recombination can’t be called into service, which makes them entangled and non-independent. Only an event of new mutation can shift a frequency from 0 to 1/N. Selection can’t. Drift can’t. Recombination can’t. Once we are in the interior of the allele-frequency space, we can navigate with the “forces”, but the forces don’t get us onto the playing field.
This is the only place where the 4 forces have any kind of parity or independence: any force can shift f to f + d, where f is a frequency such that 0 < f < 1, and d is an infinitesimal. This is what makes a force a force. By contrast, if we are not in the interior— if we are sitting on an axis waiting for a new mutation—, then selection, drift and recombination can’t be called into service, which makes them entangled and non-independent. Only an event of new mutation can shift a frequency from 0 to 1/N. Selection can’t. Drift can’t. Recombination can’t. Once we are in the interior of the allele-frequency space, we can navigate with the “forces”, but the forces don’t get us onto the playing field.
Therefore, to the extent that evolution depends on new mutations, the forces view fails to provide a sufficient description of causation.
But wait, you will object, isn’t mutation a force? Yes, there is a force in population genetics named “mutation”, but this doesn’t necessarily cover all the implications of the biological process also called “mutation”. When the biological process of mutation results in the introduction of a new allele, this is a point process, not a mass-action process. When a new allele is introduced, the system doesn’t flow off of an axis— it jumps discretely. This action does not fit how a force is supposed to act. We can’t ignore that and call the introduction process a “force” just because “new mutation” and “mutation pressure” both use the word “mutation”. If we did that, we would be mixing apples and oranges.
Failure to include a discrete novelty-introducing process is a fatal flaw in the forces theory of population-genetic causation. I do not think there is any hope for fixing it. Note that most people who dislike the “forces” view complain that it is reductionistic and makes too grand a claim for population genetics, ignoring other fields. I’m saying something completely different: the “forces” view is inadequate within population genetics.
How could such a fundamentally flawed view have become so popular? The “forces” view originated, not as a conceptual error, but as a tendentiously anti-mutational theory of evolution. The architects of the Modern Synthesis conceived of “evolution” as taking place in the interior of an allele-frequency space, because they believed (and argued explicitly) that the dynamics of new mutations could be ignored. They were determined to convince the world that evolution was the kind of smooth selection-governed mass-action process that Darwin imagined, and not the “lucky mutations” process suggested by the early geneticists, whom they dismissed as “mutationists”. To support this view, they relied on hand-waving arguments about how selection never waits for a new mutation, every species has a “gene pool” that “maintains” abundant variation, and so on. They engineered a theory that excluded mutationist thinking, and that is precisely why the theory must be judged a failure today. The wrongness of their beliefs about the role of mutation in evolution is exposed in The shift to mutationism is documented in our language. Though mistaken in their thinking about evolution, Mayr, et al were very good at rhetoric, and gained many followers with their writings. We are still trying to dig our way out from under the false sense of consensus that they created, known today as “the Modern Synthesis”.
The next problem is historical
Darwin articulated a clear but informal description of one of those forces, selection (including natural and sexual selection), whose central role in the evolution of complex phenotypic traits is universally accepted, and for which an elaborate formal theory in terms of changing genotype frequencies now exists.
Yes, there is an elaborate theory today. No, Darwin did not articulate a force of selection like the population-genetic force. If you believe in blending and fluctuation, like Darwin did, you can’t even conceive of selection this way, because there is no constancy, no heritable state to which you can attach a frequency (or a selection coefficient). The early Mendelians, faced with the challenge of understanding adaptation in a context of discrete inheritance that Darwin and his followers rejected, re-conceptualized selection as a frequency-shifting force, a process that increases one variant at the expense of another. Historian Jean Gayon refers to this as “the most important event in the history of Darwinism: the Mendelian reconstruction of the principle of selection” (p. 289; click for Google books version).
Let’s move on to the next 2 sentences.
The remaining three evolutionary forces, however, are non-adaptive in the sense that they are not the function of the fitness properties of individuals: mutation (broadly including insertions, deletions, and duplications) is the fundamental source of variation on which natural selection acts; recombination (including crossing-over and gene conversion) assorts variation within and among chromosomes; and random genetic drift insures that gene frequencies deviate a bit from generation to generation independently of other forces.
Given the century of theoretical and empirical work devoted to the study of evolution, the only logical conclusion is that these four broad classes of mechanisms are, in fact, the only fundamental forces of evolution.
Even if we were to accept the forces analogy, it would not be correct to say that a century of study has failed to reveal any new mechanisms. What about lateral gene transfer (LGT) and endosymbiogenesis? Is Lynch going to include those in “mutation”? That would be cheating, because the mutation concept in classical population genetics never involved transferring genes (or membrane-bound compartments) from one species to another. Perhaps more poignantly for Lynch’s argument, BGC (biased gene conversion) is now accepted as a population-genetic force, a means of fixation important in nature. Decades ago, Gabby Dover argued along similar lines for a new population-genetic force of “molecular drive”, and was scorned. BGC, like Dover’s process of “molecular drive”, is not the same thing as recombination. The recombination operation [1] takes A1_B1 and A2_B2 and makes A1_B2 and A2_B1. The BGC operation propagates an allele from one chromosome to another, changing A1_B1 and A2_B2 into A1_B1 and A2_B1. Not the same. Gene conversion is a kind of non-Mendelian inheritance (it violates the classic Mendelian rule that hereditary factors combine and segregate in reproduction without mixing or transforming each other). 
Don’t be confused by the fact that the cellular mechanisms that produce gene conversion and BGC overlap with those producing crossing-over. The cellular mechanisms that produce BGC and recombination also produce some mutations, but we don’t conclude from this that mutation, BGC and recombination are all the same thing. We really should call BGC a new form of selection, but BGC diverges so wildly from the teleology and purpose with which Darwinians have invested “selection”, we all tend to think of BGC as something more like mutation or recombination, even though technically, it is a cousin of selection (operating, not in the arena of organismal reproduction, but in a new and very limited arena for genic reproduction opened up by the potential for gene conversion).
Using the same old language sloppily and using the same old theory precisely are two completely different things. Lynch’s argument sounds very rigorous if it really means that no new forces have been found and no old forces have been questioned. But if it really just means “The words ‘mutation’, ‘selection’, ‘recombination’, and ‘drift’ have been stretched to cover all subsequent phenomenology”, then that is quite a different argument.
I also find the second sentence impossible to reconcile. As explained in a previous blog (Theory vs. Theory), there is a world of difference between, on the one hand, a substantive falsifiable theory, and on the other, a body of theory in the sense of a set of mathematical abstractions specific to some field, consisting merely of formalisms not falsifiable by evidence. We can call these two theoryC (concrete, conjectural) and theoryA (abstract, analytical). Theoretical population genetics is a body of theoryA. It is not a theoryC. It cannot be rejected by facts. The theoryC of evolution promoted by Mayr and Dobzhansky has been rejected. Dobzhansky thought of his theoryC as being based on theoryA, and of course a theoryC typically makes reference to theoryA, but they are not the same.
I suppose Lynch is implying that, if the theoryA of population genetics from 1930 were insufficient, then over time we would have found some phenomenon that we could not explain, that gave rise to a new branch of theory.
I agree with that, partly, but it doesn’t work in Lynch’s favor. At some point, someone created a model with an LGT or a BGC operator, and at that point, we had formalized a mechanism (part of population-genetics theoryA) that had not been formalized before. QED, Lynch’s argument is wrong.
And this does not even address the innovation at higher levels of theory. Important theoretical developments in evolutionary genetics after the classic period include the theory of kin selection (Hamilton, 1964), origin-fixation models (King & Jukes, 1969; Kimura & Maruyama, 1969), evolutionary game theory (Maynard Smith and Price, 1973), the coalescent (Kingman, 1982), and the multivariate generalization of quantitative genetics (Lande and Arnold, 1983) (the list is from a recent paper). I think “adaptive dynamics” might go on that list as well. These are just the changes in abstract theoryA. Our views of how evolution actually works in nature have changed profoundly. Apropos, note the status of origin-fixation models, which treat evolution as a 2-step process of origination (by mutation) and fixation (by selection or drift). They are widely used today, and considered fundamental, but were not invented until 1969, because they ran contrary to Modern Synthesis thinking (see The surprising case of origin-fixation models and The shift to mutationism is documented in our language).
Last sentence:
Their relative intensity, directionality, and variation over time define the way in which evolution proceeds in a particular context.
As noted, drift and recombination have magnitude but not direction.
I’ll end by pointing out that there are really 2 parts to Lynch’s argument. One part is to present a characterization of population genetics. The second part is to make some claim of sufficiency on behalf of population genetics, e.g., that it has an exclusive claim to being the language of causation in evolutionary theory. I am arguing above that the characterization of population genetics using the “forces” theory is insufficient. I haven’t really addressed whether, given a better characterization of population genetics, there would be some hope that it would suffice as a mechanistic theory of evolution. I don’t think it can, for reasons discussed by West-Eberhard.
Footnotes
[1] In molecular biology, “recombination” means “crossing over”. In evolutionary genetics, “recombination” is often used quite broadly to include crossing-over, chromosome assortment, and sexual mixis, though these are completely distinct processes in terms of biological mechanisms. What they have in common is that, relative to a generic theory of discrete inheritance of constant hereditary factors, they create new combinations out of existing factors.

Leave a comment