Some of you may have noticed a recent exchange in Nature on the question of whether evolutionary biology needs a re-think. The online article does not make clear the alignments of the listed authors, but those arguing in favor of a re-think are:
- Kevin Laland, Tobias Uller, Marc Feldman, Kim Sterelny, Gerd B. Müller, Armin Moczek, Eva Jablonka, and John Odling-Smee
and those arguing against are:
- Gregory A. Wray, Hopi E. Hoekstra, Douglas J. Futuyma, Richard E. Lenski, Trudy F. C. Mackay, Dolph Schluter and Joan E. Strassmann
I was a bit surprised that they didn’t get people who actually disagree about science, like Mike Lynch and Sean Carroll. Instead, the debate takes place between participants who disagree on the meta-scientific question of whether the field needs a re-think. What is each side saying?
The reform position
Arguing in favor of a re-think, the reformers want to broaden our perspective from the narrow confines of the “mainstream evolutionary theory” or “standard evolutionary theory” (SET), which focuses on selection and gene frequencies. As a broader and yet “unified” alternative, they offer “the extended evolutionary synthesis” (EES).
EES invokes “drivers” that, the reformers say, “cannot be reduced to genes”, and that “must be woven into the very fabric of evolutionary theory.” SET “fails to capture the full gamut of processes that direct evolution. Missing pieces include how physical development influences the generation of variation (developmental bias); how the environment directly shapes organisms’ traits (plasticity); how organisms modify environments (niche construction); and how organisms transmit more than genes across generations (extra-genetic inheritance). For SET, these phenomena are just outcomes of evolution. For the EES, they are also causes.”
The reformers go on to describe each of these causes in more detail, citing limited evidence.
They begin with developmental bias: “Rather than selection being free to traverse across any physical possibility, it is guided along specific routes opened up by the processes of development.”

The reformers summarize by saying that these results “demonstrate that development is a direct cause of why and how adaptation and speciation occur, and of the rates and patterns of evolutionary change.” And they repeat the idea that researchers are limited by SET, and that EES is a broader alternative. They stress that it is “no longer a protest movement”, but a “credible” framework with increasing support.
All is well
According to the “all is well” side, we already have an extensible framework in the Modern Synthesis (MS), which continues to be modified and expanded. The “all is well” side (1) concedes the legitimacy of every non-traditional mode of causation invoked by the “reform” side, while at the same time (2) claiming that there is no novelty in the EES position, which is just new words for old ideas, and (3) claiming that the advocates of EES haven’t made their case. Each of the topics mentioned by the reformers “already receive their due in current evolutionary theory.” Some of the topics date back to Darwin— his studies of earthworms were an early work on niche construction. The non-traditional causes of EES are interesting, to be sure, but “none of the phenomena championed by Laland and colleagues are neglected in evolutionary biology. Like all ideas, however, they need to prove their value in the marketplace of rigorous theory, empirical results and critical discussion.”
In subtle and not-so-subtle ways, the “all is well” advocates repeatedly tell the reformers to shut up and get back to work already (e.g., “advocacy can only take an idea so far”). In a final slapdown, they mention how Darwin worked for 40 years on earthworms before publishing, with the clear implication that the EES advocates should get back to work instead of blowing smoke.
Why these debates are so lame

With all due respect to the participants, this was largely a waste of time. The format was inadequate and the issue poorly framed. It is not the first time that a debate on evolutionary theory has come up short. This summer there was a thread on LinkedIn [1] that showed some of the same flaws and was tainted by the “emotional, even hostile, reaction” described by Laland, et al. What is going on in these futile debates?
Problem 1: meta-science is not science
Let us say that there is a large set of conjectural theories T about possible mechanisms or processes or phenomena in evolutionary biology, and there is a function E(T) that represents the current distribution of effort or attention given to those topics. The two sides do not disagree about what gets included in T. Instead, the reform side is arguing that E() is far from ideal, and the “all is well” side is disagreeing. If the “all is well” side is right, we just need to let the process of science work, and everything will come out right.
Really? Is the playing field level? Do reformers with important new results that build on EES themes get invited to give keynote addresses as often as Wray, MacKay, Lenski, Hoekstra, et al? [2] If I submit a grant proposal or a manuscript focused on non-traditional causes, does it have the same chance of success? What if the reviewers, citing the arguments of Wray, et al, say that EES is just a re-wording of old ideas with insufficient evidence to give it more attention than it has received already? Do Wray, et al., who claim to encourage EES work, realize how discouraging (and even disparaging) they are?
I could ask a lot of similar questions, but the point is that these are questions about how institutionalized science works, not about how evolution works. They can be answered only with sociological research that the participants have not done. These people are evolutionary biologists. Let them debate the science, not meta-science.
Problem 2: research programs are not theories
Setting aside meta-scientific arguments, we can focus on evolutionary theory. But what exactly is “evolutionary theory” for these participants?

The lack of a clear and legitimate conception is most obvious in the “all is well” side. Darwin studied earthworms, which shape their environment, an example cited today (e.g., on wikipedia) as niche construction. Does this mean that Darwin understood niche construction, and worked it into his understanding of evolutionary causation? No, absolutely not! Yet the negative side repeatedly defends “evolutionary theory” by pointing to scientists churning out data and papers, and saying “all is well”.
The fact that people are studying a phenomenon does not mean that the phenomenon is consistent with their beliefs or their causal theories. Humans have a notorious ability to be blind to unimagined possibilities, and to maintain contradictory ideas.
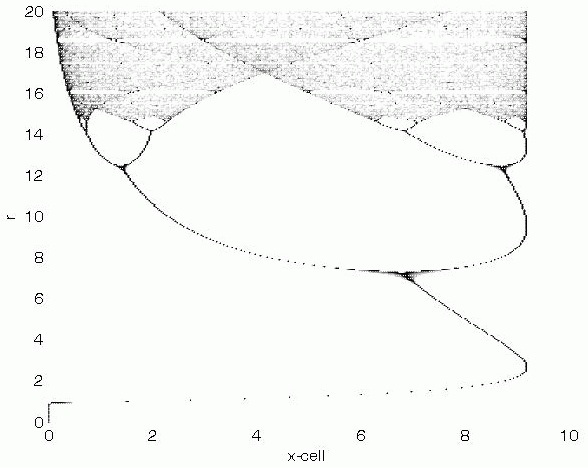
For instance, predator-prey dynamics were studied for decades before mainstream scientists realized that chaotic behavior, relevant in nature, emerges from familiar equations that had been used for years. It wasn’t that they lacked the evidence. They lacked the imagination and they lacked the concepts. John Maynard Smith, in a humorous series of anecdotes about things that he didn’t quite discover, once told me that he stumbled upon chaotic behavior of predator-prey dynamics while working out numeric examples, but didn’t know what to make of it and moved on (he also almost-discovered the neutral theory). As Winston Churchill said, “Men occasionally stumble over the truth, but most of them pick themselves up and hurry off as if nothing ever happened.” How often are evolutionary biologists stumbling over evidence of the evolutionary importance of developmental bias or plasticity, and simply ignoring it— or worse, misinterpreting it— because we lack the theoretical context for understanding it?
Problem 3: The Old Bait and Switch
As soon as we decide to focus on theory (rather than research programs or schools of thought), we run into a cryptic but important distinction:
- theoryC (concrete, conjectural), a grand hypothesis or conjecture to account for observed phenomena, as in “prion theory of disease” or “Lamarck’s theory of evolution”, “exon theory of genes”;
- theoryA (abstract, analytical), the body of abstract principles relevant to some discipline, methodology or problem area, as in “music theory”, “population genetics theory”, “theory of stochastic processes”
I’ve gone over this in a previous blog, so I won’t dwell on it here. Because theoryA and theoryC mean different things, and particularly because theoriesC are falsifiable while theoryA is not subject to empirical refutation, it is deeply problematic that the MS (Modern Synthesis) is nearly always defended today as though it were an extensible body of abstractions with no substantive content. Fisher, Mayr, and Simpson roll over in their graves every time someone claims that the MS incorporates the Neutral Theory [3].
The danger of ignoring the substantive content of the MS can be illustrated with the case of origin-fixation models, which characterize the rate of evolution as the product of (1) the rate at which new mutations enter a population, and (2) a probability of fixation, e.g., K = 4Nus. Such models are widely used today in practical applications of phylogenetics, comparative genomics, and molecular evolution, and in emerging theories of adaptation. A probability of fixation was first derived by Haldane in 1922. On this basis, we might say that origin-fixation models build on classic work and are part of the MS, or at least, are consistent with it. After all, it’s just a bunch of population-genetics equations.
In reality, origin-fixation models do not fit with the MS logically or historically (see McCandlish and Stoltzfus). They did not emerge until 1969, in response to the molecular revolution. The problem isn’t that origin-fixation models are hard to construct, but that they evoke the mutationist theoryC advocated by early geneticists such as Bateson, de Vries, Punnett and Morgan, who said “evolution has taken place by the incorporation into the race of those mutations that are beneficial to the life and reproduction of the organism” (p. 194 of 1916). In this “lucky mutant” view, selection acts as a stochastic filter, and events of mutation play a dispositional role in determining the rate and direction of evolution. The architects of the MS rejected this way of thinking, which they ridiculed as “mutationism”, opting for a “gene pool” view in which evolution consists of continuous shifts from one optimal multi-locus distribution of allele frequencies to a new optimum (this is the between-the-lines story in Provine, 1971; we’ve discussed it at length in the “mutationism myth” blog series, and in Stoltzfus & Cable, 2014).
Has the MS evolved to include mutationism? According to “all is well”, the MS continues to be updated an improved.
IMHO, this whole “the MS has evolved” line of reasoning is thin camouflage for a bait-and-switch argument that has played a major role in subverting evolutionary discourse over the past 30 years. Whether or not we allow theories to evolve, we also must recognize substantive distinctions [4]. Our history books present the mutationist view of Bateson, et al as an errant, non-Darwinian view that was rejected on empirical and theoretical grounds. The architects of the MS saw the lucky mutant view as a distinct non-Darwinian theoryC of evolution, one that could be rejected along with “saltationism” (the idea that evolution is not strictly gradual but includes steps or jumps). If mainstream thinking now welcomes non-Darwinian ideas of mutationism and saltationism, you can call that a case of “the Synthesis evolving”, but this does not relieve us of the obligation to say that Fisher, Mayr and Dobzhansky were wrong, and that we have not only added new ideas to evolutionary thinking, but we have returned to explore ideas that they rejected.
The argument that the MS is merely a framework that adapts to new findings is a rhetorical device with little probative value. It allows defenders of the MS to claim that the MS is still in effect without being clear about what the MS was, or is. In reality, the only reason that the MS seems like a “framework” is that this is all that is left of it— the flesh has rotted away, leaving only the hard bits.  The connective tissue that held together the original MS was its allegiance to Darwinian doctrines of gradualism, the creativity of selection, and the randomness of mutation. These doctrines, inherited from Darwin and his early followers, drove the commitment to gradualism, the rejection of the more ecumenical views of early geneticists, and the development of an elaborate and speculative “gene pool” view of population genetics that would be rejected today if anyone actually remembered what it was.
The connective tissue that held together the original MS was its allegiance to Darwinian doctrines of gradualism, the creativity of selection, and the randomness of mutation. These doctrines, inherited from Darwin and his early followers, drove the commitment to gradualism, the rejection of the more ecumenical views of early geneticists, and the development of an elaborate and speculative “gene pool” view of population genetics that would be rejected today if anyone actually remembered what it was.
Note that Laland, et al accept the idea that the MS is a framework. By doing so, IMHO they have given up their advantage and made it difficult to argue for reform.
Nevertheless, I also agree with their criticism of the framework. One of the persisting bits of this framework is the idea that evolution is a theory of population-genetic “forces” (see Ch. 1 of Sober’s The Nature of Selection). The claim that evolution, including macroevolution, can be reduced to the operation of a small set of forces is understood to be one of the triumphs of the MS. This view is still taught in textbooks and defended, though we have shown that it is formally inadequate to depict the role of mutation in evolution. Unfortunately, while Laland, et al criticize this view, the “all is well” side does not include the scientists who defend it (see below).
Problem 4: Reflexively Darwinian view of theory evolution
Finally, defenders of orthodoxy adopt a perspective on theory-evolution that makes arguments for reform very difficult: they assume that the evolution of ideas is merely a series of shifts in emphasis among pre-existing ideas.
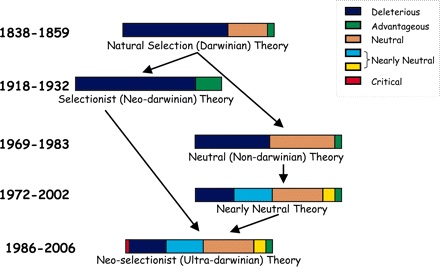
The “all is well” side apparently believes that evolutionary biology has produced no new ideas, only the re-labeling of old ideas going back to Charles Darwin. Our only task as scientists is to do the research needed to establish their relative importance.
This is a depressing and depauperate view. More importantly, it is a distortion of history. When chaotic dynamics were discovered, this was a new idea. It was not a major revolution, but nonetheless it was a distinct shift. Of course, we could take my example of chaotic dynamics, and put it in the vague category of “randomness”, then say that chaotic dynamics is merely a manifestation of the debate over “randomness” that has been going on for 150 years. What good does that do, really?
I’m not saying we should dismiss broad categories and big ideas like chance vs. determinism, externalism vs. internalism, etc. However, there are genuinely new ideas in science and— equivalently for practical purposes— old ideas that are so obscure that they are new to the vast majority of scientists. Lynch’s thesis for genome complexity is a genuinely new idea that has emerged just in the last 10 years. Another recent example would be the recognition of biased gene conversion as a population-genetic cause distinct from selection and mutation. Though the idea has been around since the early 1970s, it is only in the last 10 years that hard-core evolutionary modelers got involved in trying to understand large-scale heterogeneity in GC content within mammalian genomes (what used to be called “isochores”), and made a reasonably convincing case for the importance of GC-biased gene conversion. From now on, I suspect, biased gene conversion is going to be listed in textbooks under the category of mechanistic causes of evolutionary change.
We need to emphasize distinct differences, as when Laland, et al say “Mathematical models of evolutionary dynamics that incorporate [an ESS cause] . . . make different predictions from those that do not”. This is the kind of statement that we need to make over and over again. Models that incorporate { chaotic dynamics, biased gene conversion, mutational hazard (Lynch), non-infinitesimal variation, . . . } make different predictions.

Leave a comment